Chapter 6 ~ Evolution
Key Concepts
After completing this chapter, you will be able to
- Explain differences in environmental conditions before and after the natural genesis of life.
- Discuss the differences between creationism and evolution as explanations of the origin of life and species.
- Describe the theory of evolution by natural selection.
- Explain the role of genetics in understanding evolution and biodiversity.
In the Beginning …
Based on geological and astronomical data, the Earth is believed to have originated by the condensation of interstellar dust about 4.5 billion years ago. The pre-life environments of the planet were vastly different from what exists today. The initial atmosphere likely resulted from volcanic out-gassing and its chemistry was dominated by hydrogen sulphide (H2S), methane (CH4), ammonia (NH3), carbon dioxide (CO2), and other gases that today exist only in trace concentrations. In contrast, the modern atmosphere has large concentrations of oxygen (O2) and nitrogen (N2).
One reason for a profound change in atmospheric chemistry was the evolution of photosynthetic organisms, which release O2 as a waste product of their autotrophic metabolism. As the concentration of O2 increased, the atmosphere changed from an environment that favoured reducing reactions (in which the reaction products have a net gain in electrons) to one in which oxidizing reactions were predominant. Hydrogen sulphide, methane, and ammonia are all reduced compounds, but in an O2-rich atmosphere, they become oxidized to sulphate (SO42–), carbon dioxide, and nitrate (NO3–), respectively. In addition, O2 can participate in photochemical reactions that produce small amounts of ozone (O3). When present in the upper atmosphere, ozone absorbs solar ultraviolet radiation and thereby shields organisms from many of the damaging effects of this kind of electromagnetic energy.
The genesis of life on Earth is thought to have occurred in a primordial aquatic environment at least 3.5 billion years ago. It is not known exactly how life first began from inanimate matter, although many biologists believe that the process was a spontaneous occurrence. In other words, the origin of life happened naturally, as a consequence of the existence of appropriate conditions of chemistry, temperature, pressure, energy, and other environmental factors.
As such, the origin of life could have happened as a series of random events occurring under suitable conditions. Some biologists, however, believe that genesis could have taken place in a more purposeful manner, under the influence of autocatalytic (self-catalyzed) reactions that favoured the synthesis and persistence of particular organic chemicals. Under those selective influences, molecules and their interrelationships became increasingly more complex and eventually developed the qualities that define the simplest forms of life: metabolism, growth, and reproduction.
The appropriate environmental conditions for the genesis of life probably included the presence of many simple organic compounds in primordial waters. It is believed that the simple organic compounds were naturally synthesized by inorganic (i.e., non-living) reactions among the ammonia, methane, hydrogen sulphide, and other compounds that were abundant in the pre-life atmosphere. These reactions were favoured because the atmosphere at that time was a high-energy environment associated with ultraviolet radiation and lightning strikes. The resulting organic compounds were deposited into the primordial ocean by rainfall, where they became progressively concentrated, especially in shallow pools on oceanic shores, where the rate of evaporation would have been high.
Modern scientists have performed simple laboratory experiments that are thought to simulate those primordial conditions. In airtight flasks, mixtures of water and gaseous CH4, NH3, and H2S are sparked by electric arcs. These experiments yield various types of hydrocarbons, amino acids (precursors of proteins), nitrogenous bases (precursors of nucleic acids), and other organic chemicals. Scientists think that something similar happened prior to the origin of life on Earth.
However, it is an enormous step from the occurrence of appropriate environmental conditions to the spontaneous genesis of living microorganisms. Scientists do not yet understand how this momentous event—the origin of the first organisms—occurred. In fact, the boundary between complex chemical systems and living organisms is somewhat arbitrary (for example, viruses exist at this boundary). Nevertheless, there is a broad consensus among scientists that microorganisms did appear in the oceans about 3.5 billion years ago (Table 6.1). Those first microorganisms were heterotrophic consumers of the rich soup of organic compounds that had accumulated in pre-biological oceans over hundreds of millions of years. The first chemoautotrophic microorganisms evolved several hundred million years later, and the first photosynthetic ones about 2.5 billion years ago.
Table 6.1. Estimated Dates of the Origins of Important Life Forms. The data represent the time of first appearance of each type of organism in the fossil record. Source: Modified from Raven and Johnson (2004).
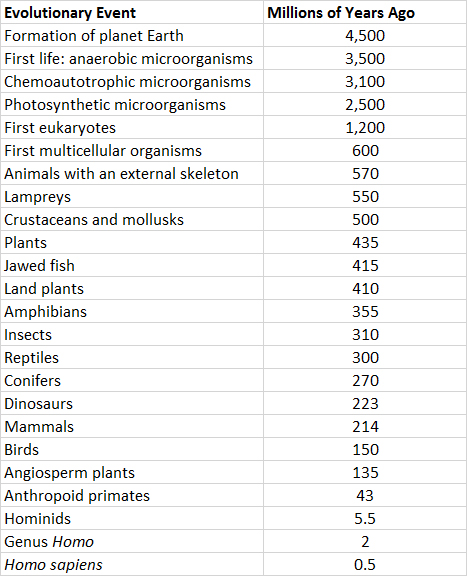
The earliest life forms were prokaryotes, which are single-celled organisms that lack an organized nucleus containing the genetic material, which was likely DNA or RNA (see In Detail 6.1). Eventually, eukaryotes (which have a nucleus bounded by a membrane) evolved from simpler prokaryotic predecessors.
More complex microorganisms, containing subcellular organelles such as mitochondria, plastids, and cilia, are thought to have evolved as a result of symbiotic associations occurring among different species. According to this theory, smaller microorganisms became encapsulated within larger ones in a mutually beneficial symbiosis (a mutualism; see Chapter 9). For example, certain smaller microorganisms may have evolved into specialized energy-processing organelles known as mitochondria. Other encapsulated microbes became specialized to capture light and to use that energy in photosynthesis—they became chloroplasts. Mitochondria and chloroplasts contain small quantities of DNA that is distinctive in character and believed to be residual from ancient times when these organelles were independent microorganisms.
Multicellular organisms were the next major category of life form to appear, in late Precambrian times (see Tables 3.1 and 6.1). The evolution and radiation of these complex organisms was driven by physiological and ecological adaptations associated with interactions of specialized cells and, eventually, organs. The first multicellular organisms were small and simple, but these eventually evolved into the larger, more complex organisms that are now prominent on Earth, including vertebrates, the phylum of animals to which humans belong.
“Progression” of Life
All species, from the smallest and simplest, such as bacteria tinier than 1 µm, to enormous blue whales exceeding 30 m in length, represent well-adapted and marvellous examples of the diversity of organisms. Moreover, in a sense modern biologists believe that all living species are similarly “advanced.” The two reasons for thinking this are: (1) all living species have had the same amount of time to evolve since the first organisms appeared, and (2) they are all exquisitely adapted to coping with the opportunities and constraints presented by the environments in which they live.
Of course, species also vary enormously in their complexity. We should, however, be careful when we use the terms “simple” and “complex” in an evolutionary context, because these concepts are difficult to precisely define. In fact, all organisms display a mixture of traits, some of which evolved in ancient times, while others are more recent adaptations. For example, almost all organisms (except some viruses) have DNA as their genetic material, so this is an ancient trait. In contrast, flight in bats and intelligence in humans represent specific adaptations that occurred relatively late in only a few evolutionary lineages.
The fossil record clearly demonstrates that, over time, there has been a progression of life forms on Earth. The first prokaryotic organisms were tiny and simple, but through evolution these led to the development of more complex eukaryotic microorganisms, and so on until large, exceedingly complex animals and plants evolved. This evolutionary pattern implies a clear temporal sequence. Nevertheless, it is important to understand that relatively complex, more recent species (including humans) do not represent the acme of evolution, nor have they inherited the Earth and its opportunities. Rather, all living species share this bountiful planet and its biosphere – the only place in the universe known to sustain life.
Image 6.1. Dinosaurs (order Dinosauria) were dominant animals on Earth for about 160 million years, but the last species became extinct 65 million years ago. We know that dinosaurs used to exist because their fossilized bones have been discovered on all continents. Modern reptiles are relatives of dinosaurs, and birds are their surviving descendants. This model of Troodon formosus, a predatory dinosaur, is located in the Museum of Nature in Ottawa. Source: B. Freedman

In Detail 6.1. A Primer on Genetics
Every organism has an individual complement of genetic information contained in the specific arrangement of nucleotides in its DNA or RNA. The following is a brief outline of the storage and translation of genetic information.DNA (deoxyribonucleic acid) carries the genetic information in almost all species. In some viruses, however, the genetic information is contained in RNA (see below). DNA, a nucleic acid, consists of linear sequences of only four nucleotides: adenine, cytosine, guanine, and thymine. The sequences are arranged as two strands, which coil as a double helix (spiral) and are held together by hydrogen bonds between complementary nucleotides: adenine with thymine, and cytosine with guanine. The genetic information is embedded in the precise sequence of the nucleotides.
RNA (ribonucleic acid) is composed of a single strand of nucleotides. In RNA, uracil substitutes for the thymine of DNA. The nucleotide sequences of RNA guide the translation of the genetic information of DNA into the structure of proteins (see below).
Chromosomes are composed of DNA and protein and they contain the genetic information of the cell. Chromosomes are self-duplicating—they create exact copies of themselves through the process of replication (see below). An exact copy is passed to each daughter cell when a cell divides. Chromosomes in body (somatic) cells of plants and animals occur as complementary pairs (homologous pairs). The number of pairs of chromosomes varies greatly among species, from one to hundreds.
Genes are specific regions of a chromosome that determine the development of a particular trait by coding for a specific protein during transcription (see below). Because chromosomes occur in pairs, the genes also are paired. Genes commonly occur in more than one form, each of which is called an allele. Often, one allele is dominant (D) and the other recessive (r). The dominant one is expressed when both alleles in a gene pair are of this type (DD), and also when both dominant and recessive alleles occur (rD or Dr). Recessive alleles are expressed only if both are of this type (rr). The condition in which both alleles are the same (DD or rr) is referred to as homozygous, while the mixed condition (rD or Dr) is heterozygous.
Replication is the biochemical process during which the nucleotide sequence of each strand of DNA is copied. Replication is necessary for cellular division to occur, because each new cell requires an identical copy of the DNA. During replication, the double helix of DNA “unzips,” which allows free nucleotides to hydrogen-bond with those in each strand, producing new but identical DNA molecules. If an error occurs during replication, the result in a change in the genetic information, which is called a mutation.
Transcription involves DNA unzipping and a complementary strand of RNA being made on one of the DNA strands, in a manner similar to replication. Then the RNA floats free and the DNA zips up again. Three types of RNA can be made: (a) ribosomal RNA (rRNA), which forms small bodies in the cytoplasm called ribosomes; (b) messenger RNA (mRNA), which transports information from DNA to the ribosome; and (c) transfer RNA (tRNA), which is described below.
Translation occurs when the mRNA, which contains information from a portion of a DNA strand, attaches to a ribosome in the cytoplasm (outside of the nucleus). There, tRNA molecules bind to specific amino acids and transport them to the mRNA in the correct sequence for the synthesis of a particular protein. (Amino acids are the building blocks of proteins. Only 20 amino acids are common, but they make up the extraordinary diversity of proteins that are found in organisms. Proteins are extremely important, mainly as structural chemicals and metabolism-regulating enzymes.) The information on the mRNA, copied from the DNA, determines the exact sequence of amino acids in a protein, and therefore determines its function.
Meiosis is important in sexual reproduction, in which two “sex” cells, one from each parent, combine to start a new life. If those cells were somatic cells, each would have the same number of chromosomes as the parent (the diploid number), and the progeny would then have double the number of the parent. However, this does not occur because sex cells are not diploid. Instead, through meiosis, the number of chromosomes in sex cells is halved (to haploid), so the progeny has the same number of chromosomes as the parent.
During meiosis, the paired chromosomes separate, with one of each pair going randomly to each daughter “sex” cell. Just before they separate, exchanges of genetic material may occur between the paired chromosomes—a phenomenon known as crossing-over. Both of these processes increase the variability of genetic information in sex cells. When the haploid sex cells (one from each parent) combine, the result is a diploid progeny. Having chromosomes from each parent, the progeny is genetically different from them, but also similar. This is how parents pass their genetic information to their offspring.
Genotype refers to the unique genetic information of individual organisms, as embodied in the nucleotide sequences of their DNA. The unique genotype of an individual is fixed (except for rare mutations). However, the collective genotypes of populations and species are quite variable, although this is restricted by the range of genetic variation among the constituent individuals.
Phenotype refers to the actual expression of an individual’s genotype in terms of its anatomical development, behaviour, and biochemistry. For example, recessive alleles, unless in a homozygous condition, are not expressed, even though they appear in the genotype.
More importantly, the expression of genetic potential is also affected by environmental conditions and other circumstances. For instance, a geranium plant, with a fixed complement of genetic information, may be relatively tall and robust if it is grown under well-watered, fertile, uncrowded conditions. However, if that same individual were grown under drier, less fertile, more competitive conditions, its productivity and appearance would be quite different. Such varying growth patterns of the same genotype represent a phenotypically “plastic” response to environmental conditions. In contrast, the flower colour of individual geraniums (which can be white, red, or pink) is fixed genetically and is not affected by their environmental conditions.
The ability of an individual to exhibit phenotypically plastic responses to environmental variations is itself genetically determined to some extent. Therefore, phenotypic plasticity reflects both genetic capability and varying expression of that capability, depending on the circumstances met during the life of an individual.
Evolution
Evolution may be simply defined as genetically based changes in populations of organisms, occurring over successive generations. Evolution is a critically important theory because it accounts for the development of existing species from progenitors that may have been unlike their descendants in form and function. The reality of evolution is widely accepted by scientists, as much so as the theory of gravity, which explains how the Earth revolves around the Sun as well as many other aspects of the organization of the universe.
Natural selection is believed to be an especially important cause of evolutionary change. In essence, natural selection predicts that individual organisms that are better adapted to coping with the opportunities or limitations of their environment will have an increased likelihood of leaving descendants. If the adaptive advantages are genetically determined, they will be passed to some of the progeny, then to subsequent generations, and so on. This process will result in evolutionary change.
Evolution can also occur in response to catastrophic influences on populations of organisms, such as a forest fire or flood. This may result in more haphazard (random) changes in the genetic structure of a population. Small populations are particularly subject to such non-selective evolutionary influences. Evolution may also occur in response to choices made by humans of desirable traits in certain species—this is known as cultural selection (or artificial selection).
It is important to understand that individual organisms do not evolve. Evolution is a process of genetic change from generation to generation, occurring in populations or higher-order groupings of organisms (such as species). This is not to say that individual organisms cannot display variable responses to environmental conditions. These responses are, however, constrained by the degree of biochemical, developmental, and behavioural flexibility that is allowed by the genetic complement of each individual (its genotype). The variable expression of the genetic information of an individual is called phenotypic plasticity, but this response to variations in environmental conditions is not evolutionary change. For evolution to occur there must be a change in the collective genetic information of a population or species.
Evolution can occur at various scales. Evolutionary biologists use the term microevolution to refer to relatively subtle changes occurring within a population or species, often within only a few generations. This may lead to the evolution of a variety, race, or subspecies. In contrast, macroevolution describes the evolution of new species or higher taxonomic groups, such as a genus, family, or class. Evolutionary biologists continue to debate and discuss the linkages of these scales of evolution. Are patterns of macroevolutionary change simply the cumulative effects of many microevolutionary changes over long periods of time? Or is macroevolution actually a result of large changes occurring over a short time, each representing a great step (or saltation) of evolution? Or does macroevolution happen in both ways?
Despite debates regarding many of its details, the theory of evolution is a unifying theme in biology. This is because evolution can be used to understand so many phenomena in nature. Evolution is used by scientists to explain both the origin of life, as well as the extraordinary changes that have occurred in organisms over the billions of years of biotic history on Earth.
Relatedness and Descent
A biological definition of species is “a group of organisms that is reproductively isolated from other such groups.” Within a species, individual organisms tend to resemble each other, but more importantly, they can breed with each other and produce fertile offspring. An inability to successfully interbreed implies reproductive isolation.
That species have evolved from earlier progenitors is a well-established theory, richly supported by evidence. Some of the most compelling lines of argument, showing evolutionary patterns of relatedness and descent, are explained in the following sections.
Patterns in the Fossil Record
A well-known example of evolution that is supported by evidence in the fossil record is that of the horse lineage. One of the earliest horse-like progenitors was Eohippus, a dog-sized creature that lived about 50 million years ago. Its foot had two fused and three separate toes. Comparison of the morphology (structure) of fossil bones suggests that Eohippus was an ancestor of Mesohippus, a larger animal that lived 35 million years ago. Its foot had three fused central toes and two free outer ones. Further evolution led to Merychippus, a somewhat larger animal living 20 million years ago that also had three fused and two free toes. Next came Pliohippus, a pony-sized animal living 10 million years ago, that had all five toes fused into a hoof. Modern horses evolved several million years ago and have a hoof like that of Pliohippus. They include the horse (Equus caballus), donkey (E. asinus), Mongolian wild horse (E. przewalskii), and zebra (E. burchelli).
Inferences from Modern Species
Modern species display many obvious similarities and dissimilarities that can be used to group them on the basis of inferred relatedness. Early studies of this sort mostly involved comparative anatomy. Research on animals relied mostly on the characteristics of bones, shells, skins, and other enduring structures, while studies of plants largely involved the anatomy of flowers and fruits. More recent studies gather a much wider range of comparative information to examine relatedness among groups of species, including information about their behaviour, ecology, proteins, and—most recently—specific base sequences of DNA. For example, studies involving DNA and blood proteins have clearly shown that humans are closely related to other great apes, such as the chimpanzee, orangutan, and gorilla. Of these, humans are most closely related to chimpanzees – in fact, the two species share about 99% of the information encoded in their DNA. These observations do not suggest that humans evolved from modern apes. Rather, the appropriate interpretation is that humans and living apes share common, ape-like ancestors.
Evolution Observed
As was just described, patterns of relatedness and descent can be inferred from comparative studies of the fossil record and of the attributes of modern species. However, it is important to understand that the evolution of a new species has never been directly observed. This is because it takes a very long time for populations of related organisms to diverge enough to become new species – perhaps thousands to hundreds of thousands of years. In spite of this, biologists have no doubt that new species have been evolving for billions of years—in fact, throughout the history of life.
Although speciation has not been observed in nature, clear examples of microevolution are known. These cases provide key evidence in support of the theory of evolution.
Industrial Melanism
One example of microevolution is that of the peppered moth (Biston betularia) of western Europe. The normal coloration of this moth is a mottled, whitish tan. During the day, the moth often rests on lichen-covered trees, where it is difficult to see against the bark. This camouflage is important to the moth’s survival because its most important predators, such as birds, hunt using vision.
About a century ago in England, it was observed that some populations of peppered moths had developed a black coloration, known as melanism. This had apparently occurred in response to changes in local tree bark, which had lost its lichen cover because of air pollution and had become blackened by the deposition of soot. In these habitat conditions the normal light-coloured moths were highly visible to predators and were at a selective disadvantage compared to darker moths. Studies showed that melanism is genetically based, and that melanistic moths occurred, but were rare, in unpolluted habitats. However, melanistic individuals became dominant in populations living in polluted habitats, representing a population-level genetic change. This famous example of microevolution, which has also been demonstrated in other species of moths, is known as “industrial melanism”.
Interestingly, air quality has greatly improved over most of western Europe in recent decades, largely due to clean-air legislation that has reduced the use of coal as a source of energy. As a result, lichens are again growing on trees and bark surfaces have less soot. These recoveries have been accompanied by the reappearance of light-coloured peppered moths in places where their populations had been dominated by melanistic ones—another evolutionary response to changing environmental conditions.
Metal-Tolerant Ecotypes
In another example of observed evolution, several plant species were found growing on sites in England and Wales that were polluted by metal-rich mine waste. Although the soil was toxic to most plants, populations of a few species were thriving. The most common species were grasses, such as bent-grass (Agrostis tenuis). Research showed that these local populations had a genetically based, physiological tolerance of the toxic metals, and that they differed in this respect from other populations of the same species growing on non-polluted sites. The locally adapted populations, referred to as “metal-tolerant ecotypes,” were found to have evolved in as few as several years after their first exposure to the toxic soil. (This example was the first one to be documented and is famous for that reason. Canadian examples of metal-tolerant ecotypes, discovered later, are described in Chapter 18.)
Image 6.2. These are metal-tolerant ecotypes of the grass Deschampsia caespitosa, growing in metal-polluted soil close to a smelter near Sudbury, Ontario. Source: B. Freedman.

Religion and Evolution
The Book of Genesis is the first book of The Bible, an ancient text that provides the written foundation for many of the beliefs of the Abrahamic religions (Judaism, Christianity, and Islam). The description of divine creation in Genesis is the oldest written explanation of the origin of life on Earth, the existence of species, and the roles and responsibilities of humans in their interactions with the natural world. However, there are some profound disagreements between fundamentalist interpretations of Genesis and aspects of the theory of evolution. As a result, some religious interests have long attacked the theory of evolution, a circumstance that greatly intensified after the publication of Charles Darwin’s ideas about the role of natural selection in evolution (see the next section).
Nevertheless, science and religion are not irreconcilable. Indeed, for many people, physical concerns belong to the domain of science, whereas spiritual ones belong to the domain of religion.
In any event, some religious groups continue to insist upon a literal interpretation of the Bible as the ultimate authority for all knowledge. In particular, creationists reject the theory of evolution in favour of a literal interpretation of Genesis. They assert that the account given in Genesis means that God created the universe and all living organisms during a six-day period, culminating with the creation of humans. Humans were created in the physical image of God and were given authority and power to freely use the resources of Earth: “And God said, let us make Man in our image, after our likeness; and let them have dominion over the fish of the sea, and over the fowl of the air, and over the cattle, and over all the Earth and over every creeping thing that creepeth over the Earth.”
Moreover, humans were instructed to increase their populations and to exploit nature: “Be fruitful, and multiply, and replenish the Earth, and subdue it.”
Note, however, that there is some controversy about the meaning of the word “replenish” in this biblical passage. Some people have interpreted it in the sense of conservation, as in to refill or restore resources as they are used. Others, however, maintain that the original meaning was “to fill up,” and, in this sense, it referred to filling the Earth with humans and their economic activities. The latter interpretation is the one that is usually accepted in modern environmental writings that examine the relationship between humans, their economy, and the natural world.
Based on their literal interpretation of Genesis and other passages in the Bible, creationists have drawn the following conclusions relevant to evolution:
- Earth and its species are not ancient because creation occurred only a few thousand years ago.
- Species are essentially immutable, having been created as entities that have not changed since their creation.
- Because species were individually created, existing species did not descend from earlier ones through evolution.
- Humans are particularly special, having been created in the Creator’s image—they are not related to or descended from any other species.
But these ideas do not accord with scientific findings, as were described on the preceding pages. In particular:
- The geological record clearly demonstrates that Earth and the solar system are extremely old, having begun to develop at least 4.5 billion years ago. Life is also ancient, having originated about 3.5 billion years ago. Earth and organisms date back much further than a few thousand years.
- The fossil record provides many examples of large changes in the characteristics of species over time, as do studies of some living species. Clearly, species are not immutable. Moreover, the existing complement of species on Earth represents only a small sample of all those that have ever lived. The fossil record demonstrates that most species that evolved during the long biological history of our planet are now extinct. Many of the extinct species, families, and even phyla have no living descendants—their entire lineage is extinct (see Chapter 7).
- The fossil record presents clear evidence of lineages among groups of organisms, indicating that living species have descended from earlier ones. In almost all cases, the progenitor species are now extinct. There are even a few examples in the fossil record of links between major groups. Perhaps the most famous of these is Archaeopteryx, a metre-long creature that lived about 150 million years ago. It had teeth and other dinosaurian characters, but it also had a feathered body and could fly. Archaeopteryx is considered to be a link between extinct dinosaurs and living birds.
- Fossil and genetic information indicate that humans are descended from earlier, now-extinct species and genera. Fossil records show that the human species (Homo sapiens) is derived from an evolutionary lineage of anthropoid apes. There are a few other surviving species in that lineage, with chimpanzees, and to a lesser degree gorillas and orangutans, being the closest living relatives of humans. All surviving members of the ape family are descended from now-extinct progenitors.
People known as scientific creationists also insist that their interpretation of Genesis is the most reliable source of knowledge about the origin and evolution of life. Scientific creationists have attempted to explain some of the discrepancies between their beliefs and current scientific understanding of evolution. For example, some of them acknowledge that geological and fossil evidence suggests that Earth and life are ancient phenomena and that most species have become extinct. Most scientific creationists also acknowledge that species have changed over time, but only through microevolution—they do not agree that macroevolution has led to the development of new species from earlier ones. By extension, scientific creationists also do not believe that humans are descended from previous species of hominids or are related to other ape-like creatures or other primates. Moreover, the theory of scientific creationism does not abandon the notion that, at one particular time in the past, God created all species that have ever lived on Earth.
Science proceeds by careful observation and hypothesis testing. But scientific creationism rests on a belief, not a testable hypothesis, concerning a literal interpretation of the Bible as representing “truth” and “knowledge”. Most predictions of scientific creationists cannot be tested by rigorous methodology, but when they can be, they are refuted by the evidence. In short, despite its name, scientific creationism is not science.
Evolution by Natural Selection
Because organisms vary in their genetics and phenotypes, they also differ in their abilities to deal successfully with stresses and opportunities in their environment. Under certain conditions, an individual with a particular phenotype (which is substantially determined by its genotype) may be relatively successful compared with others having different genotypes and phenotypes.
In the sense meant here, the “success” of an individual means successful reproduction – having progeny that themselves go on to reproduce successfully. This is also referred to as fitness, or the proportionate genetic contribution made by an individual to all of the progeny in its population. A central tenet of evolutionary theory is that individuals maximize their fitness by optimizing the degree to which their own genetic attributes will influence future generations of their species.
Biologists believe that evolution proceeds mainly by natural selection, which operates when genetically based variation exists among individuals within a population, so that some of them are better adapted to deal with the prevailing environmental conditions. On average, the more-fit organisms have greater success in reproduction, and so have a disproportionate influence on the evolution of subsequent generations.
The theory of evolution by natural selection is perhaps the greatest unifying concept in modern biology, as it gives context to virtually all aspects of the study of life. This theory was co-announced publicly in 1858 by two English naturalists: Charles Darwin (1809–1882) and Alfred Russel Wallace (1823–1913). Darwin, however, had been working on aspects of the theory for about 20 years prior to its publication, and he had collected detailed evidence in support of natural selection as a mechanism of evolution. Darwin’s copious evidence was marshalled in the famous book, On the Origin of Species by Means of Natural Selection, published in 1859. Because of this book, Darwin has become more closely linked than Wallace to the theory of evolution by natural selection. Darwin is also the more famous of the two scientists, largely because of his great contributions toward understanding the mechanisms of evolution. Perhaps the most influential biologist of all time, Darwin undertook an astonishingly broad range of research projects on a great variety of species and biological topics.
In his Origin of Species, Darwin summarized natural selection in the following way: “Can we doubt … that individuals having any advantage, however slight, over others, would have the best chance of surviving and of procreating their kind? On the other hand, we may feel sure that any variation in the least degree injurious would be rigidly destroyed. This preservation of favourable variations, I call Natural Selection.”
In an unpublished essay that Wallace sent to Darwin for review in 1858, natural selection was expressed in a rather similar fashion: “The life of wild animals is a struggle for existence … in which the weakest … must always succumb … giving rise to successive variations departing further and further from the original type.”
The theory of Darwin and Wallace was based on the following line of reasoning:
- It is known that the fecundity of all species is high enough that they could easily overpopulate their habitats, yet this does not generally happen.
- It is also known that the resources that species need to sustain themselves are limited, particularly in relatively stable habitats.
- Therefore, in view of potential population growth and limited resources, there must be intense competition among individuals of each species for access to the necessities of life. Only some individuals manage to survive this struggle for existence and reproduce.
- Because individuals within a species are different from each other, and much of this variation is heritable, it is reasonable to suggest that survival in the struggle for existence is partly influenced by genetically determined differences in abilities.
- Individuals that are more capable will have a better chance of surviving and reproducing, and their genetically based attributes will be disproportionately represented in future generations.
- Over long periods of time, this process of natural selection will lead to evolutionary changes within populations, and eventually to the evolution of new species.
When it was first presented publicly in 1858, the theory of evolution by natural selection created a sensation among scientists and also within society. The excitement and controversy occurred largely because the theory provided the first convincing body of evidence in support of the following three notions: (a) evolution occurs, (b) it proceeds under natural influences, and (c) it has resulted in the great diversity of living species.
This was a radically different view from that of creationism, which was the prevailing explanation of the origin of life and species in the mid-nineteenth century. Interestingly, Darwin’s writings did not directly challenge the existence of a divine Creator. He mostly discussed the causes of change in species over time, and did not directly suggest that the initial ancestors had not been created by God. Modern theories about the spontaneous genesis of life on Earth are based on relatively sophisticated science that was unknown to Darwin. Nor did he know of the mechanisms of genetics and the inheritance of traits.
Modern extensions of the theory of evolution by natural selection suggest that new species evolve from progenitors (a process known as speciation). This is thought to happen when populations become isolated by intervening barriers such as a mountain range, extensive glaciers, or other inhospitable discontinuities. Isolation is important in speciation because it reduces or eliminates genetic interchanges, and thereby allows differentiation to proceed more effectively. Isolated populations that experience different environmental conditions are subject to differing selection pressures and can evolve in dissimilar ways. Eventually, there may be enough evolutionary change that the populations can no longer successfully interbreed, even if they become spatially reunited. At that point, the populations have achieved reproductive isolation, and so have become closely related but different species.
Speciation is also thought to occur in a more linear fashion, as when progenitor species gradually evolve over time in response to changes in environmental conditions. Eventually, the ancestral species may become extinct, but new ones evolved from the progenitor lineage may survive to continue the evolutionary chain.
The Importance of Genetics
Knowledge of genetics in Darwin’s time was based on a highly incomplete understanding of how an organism’s traits are passed to its offspring. One popular theory, the “inheritance of acquired traits”, was based on the observation that the morphology, behaviour, and/or biochemistry of individual organisms could vary in response to environmental change. According to the theory, these plastic responses to environmental conditions could be passed along to an individual’s progeny. For example, during periods of drought or intense competition for food, individual short-necked ancestors of giraffes might have stretched their necks as far as they could to reach scarce foliage higher up in trees, resulting in the development of a longer neck. The long-neckedness would have been passed to the giraffe’s progeny, who developed it still further. Eventually, populations developed the familiar long neck of modern giraffes.
However, natural selection suggests a different mechanism of this evolutionary change: within populations of short-necked giraffes there existed a genetically determined variation in neck length among individuals. Because long-necked giraffes were better able to find food, they were more likely to survive and reproduce. This meant that more of the next generation had the long-necked trait, and this anatomical feature became increasingly prominent in the evolving population.
Modern observations and experiments have shown that “acquired traits” are just a manifestation of phenotypic plasticity. There is no evidence that they can become genetically fixed in an individual and passed along to its offspring. In contrast, the science of genetics has provided convincing evidence in support of the theory of evolution by natural selection. The biochemical mechanisms that determine the genotype of an individual organism and how some of its characteristics are passed to progeny have been discovered. The subject matter is rather complicated and cannot be dealt with in much depth here. It is, however, useful to examine the key experiments that first suggested the existence of genes.
This research was conducted by Gregor Mendel (1822–1884), an Austrian scientist (and monk) who developed important ideas about inheritance through breeding experiments with the garden pea (Pisum sativum). Mendel was interested in producing pea hybrids, which involves crossing two parent plants, each having distinctive traits. Prior research had shown that certain traits were fixed in cultivated varieties of peas, including flower colour (white or purple) and whether the seeds have a wrinkled or smooth coat. In total, Mendel worked with 32 traits of this sort. Pea flowers are bisexual, containing both female (pistil, containing the ovules) and male (anthers, containing pollen) parts. These are compatible within the same individual, so self-fertilization can occur. However, Mendel experimented by cross-fertilizing selected parents, producing known hybridizations.
In each experiment, Mendel cross-bred two inbred varieties in which certain traits “bred true” (they were homozygous, such as for a white or purple flower colour). The progeny (first generation) were all the same: they all had purple flowers. However, crosses between the first-generation plants yielded a ratio of about three purple flowers to one white flower in the second generation. This fits the prediction for two-generation crosses between two homozygous lines, as follows:
- Represent the original purple variety as AA. This trait is dominant over the white flower trait (called recessive).
- Represent the original white variety as aa.
- When the two plants are crossed, the first-generation progeny all have purple flowers but are heterozygous (Aa).
- A cross between the first-generation plants yields four possible outcomes: AA, Aa, aA, and aa. Each is equally probable. Because A is dominant to a, the AA, Aa, and aA progeny all have purple flowers. Only aa has white flowers. Therefore, the expected ratio of purple to white among the second-generation progeny is 3:1.
The most important conclusion to emerge from Mendel’s work was that the inheritance of genetic information occurs in a “particulate” form (which we now refer to as genes), often involving dominant and recessive alleles. Inheritance is not a blended condition—in the example just described, a cross of purple- and white-flowered pea plants does not yield progeny of an intermediate colour. Therefore, flower colour and many other traits are discrete units that remain intact during inheritance and either are, or are not, expressed in progeny.
Mendel first published his results in 1865 in a relatively obscure journal. As a result, the work was unknown to the mainstream of science for many years. However, Mendel’s work was re-discovered and republished in 1900 and it quickly became the basis of modern theories of genetic inheritance.
Mendel’s work and the subsequent flourishing of the science of genetics have been extremely important in biology and in the development of the modern theory of evolution. This is because genetics allows a rational explanation of inheritance as a mechanism by which genetically fixed traits can be passed along to offspring. Subsequent research has found that new genotypes can arise through various mechanisms, such as hybridization, polyploidism (a spontaneous increase in the number of chromosomes), and mutations. Genetic variation is, of course, the menu of possibilities from which natural selection can choose so that adaptive evolution can occur.
It is important to recognize that much genetic information in an individual does not appear to code for functional enzymes or other proteins, and so does not code for traits that could be selected for or against. Because of its neutrality with respect to natural selection, this type of genetic material is sometimes referred to as “junk DNA”. However, we may be ignorant of other roles that so-called junk DNA may play in the functioning of the genome.
Environmental Issues 6.1. Genetically Modified Organisms
Genetically modified organisms (GMOs) are a highly controversial topic. But what, exactly, is meant by the term?Strictly speaking, GMOs are organisms whose genotype has been influenced by human intervention. But people have been doing this for an extremely long time. As early as about 10,000 years ago, when people first began to cultivate other species as crops, they selectively bred individual plants and animals that had favourable traits (see Chapter 10 for an explanation of socio-cultural evolution, including the early development of agriculture). This “artificial selection” rapidly led to the evolution of crop varieties that were more responsive to management and had greater yields than their wild progenitors. In this sense, almost all domesticated species of plants, animals, and microorganisms that are cultivated as sources of food, material, or energy are “genetically modified organisms.” They were produced using conventional methods of selective breeding, a process that is not very controversial.
More recently, however, new techniques in biotechnology, specifically in molecular biology, have been used to create novel genetic modifications of organisms. These techniques allow biologists to selectively insert portions of the DNA of one species into the genome of another species. This is a fundamentally different kind of genetic modification than selective breeding, and it should more properly be referred to as transgenic modification, or as recombinant bioengineering. There are potential benefits to this kind of genetic modification of crop species, including the development of varieties that are resistant to diseases or pests and that require less fertilizer or pesticide. In spite of these seeming benefits, there is controversy over transgenic biotechnology and the commercial use of GMOs, largely because of the following issues:
- Should scientists be interfering with the very foundation of life—the genetics of species—by using methods of genetic “engineering” that do not normally exist in nature?
- Do novel, transgenic organisms represent “new” varieties of designed and manufactured life that are appropriate for patenting and use for commercial gain? (In fact, various legal rulings have stated that this can be done, and some transgenic crops have become extremely profitable to owners of the patents.)
- Are important ecological risks associated with the cultivation of transgenic organisms? Because many biological and ecological unknowns are associated with this practice, “surprises” may follow from the release of these organisms into the environment, including unanticipated damage to crops, wild species, and natural ecosystems.
These are contentious and precautionary issues, and the controversy is not resolved. In some cases, illegal releases of GMO products have been made by private interests, a circumstance that reflects weakness in the regulatory control mechanisms (Clapp, 2008). Nevertheless, some GMO products have been widely commercialized and are now routinely used. For instance, transgenic GMO varieties of soybean and canola have been developed to be resistant to glyphosate, which allows this herbicide to be used on those crops. This practice results in benefits to farmers from reduced costs of energy and machinery needed to control weeds. In a similar vein, transgenic varieties of maize (corn) have been developed that contain modified DNA of the insecticidal bacterium Bacillus thuringiensis. This provides resistance to important insect pests and allows farmers to use less insecticide. These and other transgenic crops are now widely cultivated in North America (although they are banned in other countries, including most of Europe and Brazil), but relatively little is known about the biological and ecological risks that may arise when their transgenic factors escape to wild plants.
Additional Mechanisms of Evolution
Although natural selection is the most important mechanism of evolution, it is not the only one. For example, artificial selection involves the deliberate breeding of plants, animals, and microorganisms to enhance certain traits that humans view as desirable. Artificial selection has obvious parallels to natural selection in that individual organisms with particular, genetically based traits experience greater success in breeding, so they become over-represented in subsequent generations. However, traits that are favoured in artificial selection may not be adaptive in the natural world. In addition, because the breeding of desired genotypes can be controlled, evolutionary change occurs much more rapidly under artificial selection than under natural selection.
For example, maize (or corn, Zea mays) is an important crop that, through artificial selection, now differs enormously from its closest wild progenitor, a Mexican grass known as teosinte (Euchlaena mexicana). Artificial selection has caused many evolutionary changes in maize. For example, the fruiting head (consisting of the cob and seeds) is much larger than in wild ancestors of maize; the seeds have different coloration; the seeds implant securely onto the cob so they do not scatter before harvesting; the ripe fruit is tightly wrapped within enclosing leaves known as bracts, again to prevent pre-harvest losses; and there are vigorous growth responses to fertilizer application, weed control, and other cultivation practices. Moreover, without the intervention of humans through cultivation, maize would likely become extinct within only a few generations. This is partly because artificial selection has rendered its seeds virtually incapable of detaching from the cob, which in any event is tightly bound in leafy bracts. Therefore, unaided seed dispersal is almost impossible.
All domesticated plant, animal, and microbial species have undergone artificial selection for desirable traits. Sometimes, however, artificial selection proceeds in bizarre directions, with the fostering of genetic traits that are viewed as desirable for aesthetic rather than practical reasons. For example, oriental breeders of pet fish have produced some amazing varieties of goldfish (Carassius auratus) and koi (a golden-coloured variety of carp, Cyprinus carpio). These varieties, often with grotesque shapes and behaviours, would be rapidly eliminated in a wild population but are prized as unusual and valuable specimens by aficionados of these aquatic pets. Similar comments could be made about curious varieties of cats, dogs, pigeons, and many kinds of horticultural plants.
Evolution can also occur through a process known as genetic drift, or random changes in the frequencies of genes occurring in small and isolated populations. Such populations often exist on islands, or they may be created through a catastrophic reduction of a larger population because of disease, disturbance, or some other factor. The relatively small genetic base of small populations is sometimes called a “bottleneck.” Subsequent evolution is based on the restricted genetic variation of only a few individuals, which may become further reduced through the effects of inbreeding (reproduction between closely related individuals, such as siblings). Given the restricted amount of genetic variation, the evolution of a small population may proceed very differently from that of a larger population.
Image 6.3. The many varieties of dog are a result of cultural selection for desired traits, but all are the same species, Canis lupus familiaris. Source: B. Freedman.

Conclusions
Earth is the only place in the universe that is definitely known to sustain life and ecosystems. It is thought that life spontaneously arose at least 3.5 billion years ago, because of the existence of environmental conditions appropriate for its genesis. Since that origin, profound changes have occurred in the morphology and functionality of organisms through a process known as evolution. Evolution may be simply defined as changes in the genetic makeup of populations and species over time (individual organisms do not evolve). Although evolution has influenced life on Earth ever since it began, there is controversy over the mechanisms of the process. Almost all biologists believe that natural selection has been the most important cause of evolutionary change, but some think that geological catastrophes (such as meteorite strikes of the planet or intense volcanic eruptions) have also had a large influence.
Questions for Review
- How has the evolution of organisms, especially those capable of photosynthesis, resulted in important changes in chemistry of the environment?
- What evidence supports the theory of evolution?
- How is natural selection a mechanism of evolution? What are other means by which evolution can occur?
- How does artificial selection result in the evolution of domesticated species?
Questions for Discussion
- Why are many biologists reluctant to describe certain species as being “more advanced” or “more highly evolved” than others?
- How might environmental conditions experienced during your lifetime have affected your own development? Relate your answer to the phenomenon of phenotypic plasticity.
- Why is knowledge of genetics important to understanding evolutionary processes?
- Do you think there is enough scientific evidence in support of the idea of spontaneous generation of life for it to replace faith-based notions of divine creation?
Exploring Issues
- You have been asked to participate in a debate about the genesis and evolution of life. What kinds of evidence would you use to support the theory that life began from inanimate matter billions of years ago? What evidence supports the theory that humans evolved from earlier ancestors that are now extinct?
References Cited and Further Reading
Bengtson, S. (ed.). 1995. Early Life on Earth. Columbia University Press, New York, NY.
Clapp, J. 2008. Illegal GMO releases and corporate responsibility: Questioning the effectiveness of voluntary measures. Ecological Economics, 66: 348-358.
Cowen, R. 2000. History of Life. 5th ed. Blackwell Scientific, London, UK.
Coyne, J.A. and H.A. Orr. 2004. Speciation. Sinauer Associates, Sunderland, MA.
Darwin, C.D. 1859. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. Murray, London, UK.
Dawkins, R. 1996. The Blind Watchmaker. WW. Norton, New York.
Eldredge, N. 2000. The Pattern of Evolution. W.H. Freeman & Co., San Francisco, CA.
Gould, S.J. (ed.). 1993. Book of Life. Hutchinson, London, UK.
Hall, B.K. and B. Hallgrímsson. 2013. Strickberger’s Evolution. 5th ed. Jones & Bartlett Learning, Boston, MA.
Horgan, J. 1991. In the Beginning . . . . Scientific American, 264: 116-25.
Klug, W.S. and M.R. Cummings. 2011. Concepts of Genetics. 10th ed. Prentice Hall, Upper Saddle River, NJ.
Lewin, R. 1991. Thread of Life: The Smithsonian Looks at Evolution. Smithsonian Books, Washington, DC.
Mayr, E. 1982. The Growth of Biological Thought. Harvard University Press, Cambridge, MA. National Research Council. 1990. The Search for Life’s Origins. National Academy Press, Washington, DC.
Numbers, R.L. 1993. The Creationists: The Evolution of Scientific Creationism. University of California Press, Berkeley, CA.
Raven, P.H., G.B. Johnson, K.A. Mason, and J. Losos. 2013. Biology. 10th ed. McGraw-Hill, Columbus, OH.
Ridley, M. 2003. Evolution. 3rd ed. Blackwell Science, Boston, MA.
Shapiro, R. 1986. Origins: A Sceptic’s Guide to the Creation of Life on Earth. Simon & Schuster, New York, NY.